The Journal of Immunology, 1998, 161: 2586-2593. @
** Arthritis and Rheumatism Branch, National
Institute of Arthritis and Musculoskeletal and Skin Diseases, *Developmental
Endocrinology Branch, National Institute of Child Health and Human Development,
and the ^Immunology Service, Clinical Center, National Institutes of
Health, Bethesda, MD 20892
| Abstract |
|---|
| Introduction |
|---|
In contrast to IL-12, IL-10 inhibits several macrophage functions. Together
with IL-4, this cytokine stimulates the development, proliferation, and
function of Ab-producing B cells and promotes IgE production (8).
IL-10 also prevents Ag-specific T cell proliferation, inhibits the
production of the principal proinflammatory cytokines IL-12, TNF- ,
and IFN-
,
and IFN- ,
and prevents endotoxin shock in mice (reviewed in 9
. In humans, initial clinical trials demonstrated that IL-10
administration ameliorated inflammatory symptoms associated with endotoxemia
(10), inflammatory bowel disease (11), and
rheumatoid arthritis (12). Overexpression of IL-10
seems to play an inappropriate immunosuppressive role, allowing
increased malignant tumor growth, as seen in melanoma (13)
and in systemic diseases with excessive production of Abs, such as
the immune complex-related manifestations of lupus erythematosus (14).
,
and prevents endotoxin shock in mice (reviewed in 9
. In humans, initial clinical trials demonstrated that IL-10
administration ameliorated inflammatory symptoms associated with endotoxemia
(10), inflammatory bowel disease (11), and
rheumatoid arthritis (12). Overexpression of IL-10
seems to play an inappropriate immunosuppressive role, allowing
increased malignant tumor growth, as seen in melanoma (13)
and in systemic diseases with excessive production of Abs, such as
the immune complex-related manifestations of lupus erythematosus (14).
Histamine, a recognized regulator of inflammation, is also known for its role in promoting allergic reactions, gastric acid secretion, and tumorigenesis. As a reflection of its importance, histamine-blocking agents, worldwide, are the largest selling drug group today. Atopic states, associated with excessive histamine production, involve a shift from Th1 to Th2 responses and increased IgE production. On the other hand, chronic gastritis with progression into peptic ulcer disease is also associated with increased histamine production in the gastric mucosa. Chronic gastritis is largely caused by infection with Helicobacter pylori, against which Th1 responses appear to have a protective role (15). Finally, suppression of Th1 responses accompanies the development of diverse tumors, while, in some tissues, histamine itself appears to promote tumorigenesis (16). Interestingly, recent evidence indicates that the use of H2 receptor antagonists increases survival of patients with gastric and colorectal cancer (17, 18).
Thus, the question arises: Does histamine directly induce or promote a Th2 shift? Theoretically, histamine, or any other mediator, could alter the Th1/Th2 balance at the level of APC, Th1 and Th2 cells, or directly on effector cells. Since IL-12 and IL-10, as noted above, have important regulatory influences on Th1 and Th2 functions, we speculated that histamine could modulate IL-12 and IL-10 production and, thus, Th1/Th2 balance. Here we report that histamine, via an H2 receptor-mediated process, potently suppressed the production of human IL-12, while it enhanced the secretion of IL-10. This may represent a mechanism through which histamine skews Th1/Th2 cytokine balance toward Th2-type dominance.
| Materials and Methods |
|---|
Flow cytometric evaluation of manipulated cell preparations was performed using directly conjugated mAbs to the following surface Ags: CD45, CD14, CD3, CD20, CD16/56 (Becton Dickinson Immunocytometry Systems (BDIS), San Jose, CA). LPS from Escherichia coli, serotype K-235 (Sigma, St. Louis, MO), was dissolved in distilled water; the mixture was sonicated for at least 3 min in a sonicating bath, and aliquots were stored at -20°C until use. After thawing, appropriate dilutions were made in RPMI 1640. Staphylococcus aureus Cowan strain 1 (SAC) was obtained from Calbiochem-Behring, (La Jolla, CA). Histamine, dimaprit, cimetidine, pyrilamine maleate (mepyramine), thioperamine maleate, and Ro 20–1724 were purchased from Research Biochemicals (Natick, MA). Anti-human IL-10 Abs and recombinant human IL-12 were obtained from R&D Systems (Minneapolis, MN).
Blood donors
Thirty-five healthy male and female volunteers between 20 and 40 years old participated in this study, which was approved by an institutional review board of the National Institutes of Health. Volunteers abstained from using any drug, including antihistamines, cyclooxygenase inhibitors, and hormones during the week before the study.
Whole blood cultures
The whole blood assay was established recently as a suitable ex vivo model by which to study cytokine production under conditions in which many of the physiologically relevant cellular interactions and natural microenvironments remain intact (19). In addition, this method offers the opportunity to induce and detect cytokine production in small blood samples and to study multiple samples under different conditions, employing the blood from the same donor, and, thereby, eliminating interdonor variability as a confounding factor. In brief, blood was drawn into sodium heparin-containing sterile blood collecting tubes (Vacutainer, Becton Dickinson, Lincoln Park, NJ). Blood was transferred to 50 ml Falcon tubes, diluted 1:5 with RPMI 1640 (supplemented with 1% glutamine and gentamicin, 50 µg/ml) with no added exogenous serum, and aliquoted (1.0 ml) into 24-well cell culture plates (Costar, Cambridge, MA). To induce cytokine production, bacterial LPS was added at 1 µg/ml final concentration, and the samples were incubated in 5% CO2 at 37°C for 18 h. Histamine and the H2 receptor agonist dimaprit were added 10 min before LPS. Histamine antagonists and the phosphodiesterase inhibitor Ro 20–1724 were added to the wells 10 min before histamine. Anti-IL-10-neutralizing Abs or recombinant human IL-12 were added simultaneously with LPS. After incubation, the blood was centrifuged, and the supernatant plasma was collected and stored in polypropylene tubes at -70°C until assayed.
PBMC, PBL, and monocyte isolation and culture
Human peripheral blood samples were obtained by leukapheresis of
volunteers at the National Institutes of Health Department of
Transfusion Medicine. PBMC were isolated by density gradient centrifugation
using Ficoll-Hypaque (Histopaque, Sigma). To obtain monocyte-enriched
preparations, total PBMC were depleted of T and B lymphocytes using
magnetic beads (Dynal, Lake Success, NY) coated with mAb specific for
the CD2 (Pan T cells) and CD19 (Pan B cells) surface Ags. Total PBMC
at 10 to 15 x 106 cells/ml were
incubated (two times, with >10 beads per target cell) and gently
mixed with the magnetic beads at 4°C for 30 min. The bead-cell
complexes were eliminated using magnetic separation. The
monocyte-enriched preparations contained 73 to 89% monocytes (n
= 4), as determined by flow cytometry (FACScan, BDIS) with
monocyte-specific FITC-conjugated anti-CD14. Lymphocyte contamination
ranged from 11 to 27% and consisted of 60 to 75% NK cells, 20 to 35%
T cells, and  3%
B cells. Lymphocytes (PBL) were obtained by depleting monocytes from
total PBMC using magnetic beads coated with a primary mAb specific for
CD14. A single step incubation and elimination was performed as
above. The PBL preparations contained 0.01%
monocytes (3 of 4 had 0%), as determined by flow cytometry.
3%
B cells. Lymphocytes (PBL) were obtained by depleting monocytes from
total PBMC using magnetic beads coated with a primary mAb specific for
CD14. A single step incubation and elimination was performed as
above. The PBL preparations contained 0.01%
monocytes (3 of 4 had 0%), as determined by flow cytometry.
PBMC were cultured at 1 x 106 cells/ml, while PBL and monocytes were cultured at 5 x 105 cells/ml. All cultures were set up in RPMI 1640 medium supplemented with 15% FCS, 1% glutamine, gentamicin 50 µg/ml at a final volume of 0.5 ml. PBMC were activated with SAC at a final dilution of 1:200, while PBL and monocytes were stimulated with LPS (1 µg/ml). All cultures were incubated in 5% CO2 at 37°C for 18 h.
Cytokine assays
IL-12 (p70 and p40) and IL-10 were measured using ELISAs employing the multiple Ab sandwich principle (Quantikine, R&D Systems). These assays specifically detect human IL-12 p70 (the biologically active heterodimer), the 40-kDa (p40) subunit of IL-12 and human IL-10, respectively. IL-12 p70 ELISA recognizes specifically the IL-12 heterodimer without cross-reactivity with the individual subunits of the dimer (p35 and p40). The detection limits of the IL-12 p40, IL-12 p70, and the high sensitivity (HS) IL-12 p70 ELISAs were 15.0, 5.0, and 0.5 pg/ml, respectively, while for the IL-10 ELISA it was 2 pg/ml. Plates were read by a microplate reader (Model 550, Bio-Rad, Richmond, CA), and absorbency was transformed to cytokine concentration (pg/ml) using a standard curve computed by Microplate Manager III, Macintosh Data Analysis Software (Bio-Rad).
| Results |
|---|
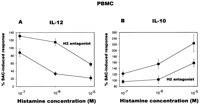
Addition of increasing concentrations of histamine to the whole blood culture,
resulted in a dose-dependent inhibition of LPS-induced IL-12 p70
production (Fig. 1![]() A).
Histamine at 10-5 M concentration resulted in >95%
inhibition of IL-12 production. Conversely, increasing concentrations
of histamine caused a dose-dependent increase of LPS-induced IL-10
production (Fig. 1
A).
Histamine at 10-5 M concentration resulted in >95%
inhibition of IL-12 production. Conversely, increasing concentrations
of histamine caused a dose-dependent increase of LPS-induced IL-10
production (Fig. 1![]() B),
yielding a substantial increase of IL-10 production at 10-7
M (186% increase) and 10-6 M (521% increase).
B),
yielding a substantial increase of IL-10 production at 10-7
M (186% increase) and 10-6 M (521% increase).
|
The addition of increasing concentrations of the H1 receptor antagonist,
pyrilamine maleate (mepyramine), and the H3 receptor antagonist, thioperamine
maleate, failed to prevent the effect of histamine (0.1 µM) on both
IL-12 and IL-10 secretion (Fig. 2![]() ,
A, C, D, and F). However, both the
inhibitory and the stimulatory effects of histamine (0.1 µM) on
IL-12 and IL-10, respectively, were antagonized by increasing
concentrations of cimetidine, an H2 receptor-blocking agent, leading
to a complete blockade of the effect at 10 µM concentration of
cimetidine (Fig. 2
,
A, C, D, and F). However, both the
inhibitory and the stimulatory effects of histamine (0.1 µM) on
IL-12 and IL-10, respectively, were antagonized by increasing
concentrations of cimetidine, an H2 receptor-blocking agent, leading
to a complete blockade of the effect at 10 µM concentration of
cimetidine (Fig. 2![]() ,
B and E). These results suggest that the effects of histamine
on these cytokines were mediated by the H2 receptor. Increasing doses
of the H2 antagonist cimetidine, identical to those used to block the
effect of histamine, without added histamine did not affect
LPS-induced IL-12 p70 or IL-10 production (n = 4; data not
shown).
,
B and E). These results suggest that the effects of histamine
on these cytokines were mediated by the H2 receptor. Increasing doses
of the H2 antagonist cimetidine, identical to those used to block the
effect of histamine, without added histamine did not affect
LPS-induced IL-12 p70 or IL-10 production (n = 4; data not
shown).
|
To demonstrate the involvement of H2 receptors in the histamine effect more
definitively, we used dimaprit, an H2 receptor agonist in the
subsequent experiments. As evident from Figure 3![]() ,
A and B, the addition of this drug induced
dose-dependent inhibition and potentiation of LPS-induced IL-12 and
IL-10 production, respectively. Thus, the effect of this drug fully
mimicked the effect of histamine on the production of these
cytokines.
,
A and B, the addition of this drug induced
dose-dependent inhibition and potentiation of LPS-induced IL-12 and
IL-10 production, respectively. Thus, the effect of this drug fully
mimicked the effect of histamine on the production of these
cytokines.
|
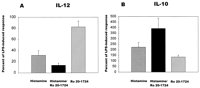
In subsequent experiments, we tested the possibility that histamine itself, without LPS, is able to induce or reduce IL-12 or IL-10 production. Addition of increasing doses of histamine (10-9-10-5 M), without LPS, failed to affect basal IL-12 production. The level of IL-12 at this concentration range of histamine was <0.5 pg/ml, similar to the negative control (no histamine), which was 0.5 ± 0.1 pg/ml (n = 4). Similarly, increasing doses of histamine (10-9-10-5 M), without LPS, failed to affect IL-10 production. The level of IL-10 at this concentration range of histamine was between 2.0 and 2.8 pg/ml, similar to the negative control (no histamine), which was 3.5 ± 1.1 pg/ml (n = 4).
The effect of histamine is independent of endogenous IL-10 production and is not affected by administration of exogenous IL-12
IL-10 has been described to inhibit the secretion of proinflammatory cytokines
from monocytes/macrophages. This prompted us to investigate the
possibility that the inhibitory effect of histamine on IL-12 production
was caused by induction of endogenous IL-10 secretion. Conversely,
since IL-12 inhibits IL-10 production, we decided to test whether
administration of exogenous IL-12 might interfere with the effect of
histamine on IL-10 production. For this purpose, we investigated the
effect of adding neutralizing Abs to IL-10 or exogenous IL-12 to
whole blood cultures stimulated with LPS in the presence of
histamine. The concentration of anti-IL-10 Abs used in our
experiments was 10 µg/ml. This concentration, according to the
manufacturer’s instructions, gives a 50% neutralizing dose (ND50)
in the presence of 5 ng/ml of rhIL-10. The mean LPS-induced IL-10
levels in our experiments were 300 ± 81.3 pg/ml (Fig. 4![]() A).
Thus, the concentrations of endogenously produced IL-10 were about
16-fold less than the concentration at which anti-IL-10 Abs express
ND50. Thus, we concluded that the concentration of
neutralizing anti-IL-10 Abs used in these experiments was sufficient
to eliminate endogenously produced IL-10. As shown in Figure 4
A).
Thus, the concentrations of endogenously produced IL-10 were about
16-fold less than the concentration at which anti-IL-10 Abs express
ND50. Thus, we concluded that the concentration of
neutralizing anti-IL-10 Abs used in these experiments was sufficient
to eliminate endogenously produced IL-10. As shown in Figure 4![]() A,
the inhibition of IL-12 induced by histamine (0.1 µM) could not be
prevented by anti-IL-10 Abs. The LPS-induced levels of IL-12 p70 in
our whole blood system usually ranged between 20 and 100 pg/ml. For
this reason, we employed 50 and 200 pg/ml of exogenous IL-12 in our
experiments. As shown in Figure 4
A,
the inhibition of IL-12 induced by histamine (0.1 µM) could not be
prevented by anti-IL-10 Abs. The LPS-induced levels of IL-12 p70 in
our whole blood system usually ranged between 20 and 100 pg/ml. For
this reason, we employed 50 and 200 pg/ml of exogenous IL-12 in our
experiments. As shown in Figure 4![]() B,
the exogenously administered IL-12 did not reverse the
histamine-induced enhancement of IL-10 production.
B,
the exogenously administered IL-12 did not reverse the
histamine-induced enhancement of IL-10 production.
|
In the preceding experiments, we observed that the effect of histamine was
mediated by H2 receptors. In many tissues, including immune cells,
the stimulation of H2 receptors results in increased cAMP production.
Phosphodiesterase is an enzyme involved in the breakdown of cAMP.
Inhibition of this enzyme results in increased levels of cAMP. We
investigated the effect of Ro 20–1724, a specific phoshodiesterase
inhibitor. In our assay, Ro 20–1724 itself (1 µM), without
histamine, induced only a slight, nonsignificant inhibition or
potentiation of LPS-induced IL-12 and IL-10 production. However, the
addition of Ro 20–1724 to cultures containing histamine induced
twofold potentiation of the inhibitory effect on IL-12 and the
enhancing effect on IL-10 production (see Fig. 5![]() ,
A and B). These observations suggest the involvement of
cAMP production in these immunoregulatory effects by histamine.
,
A and B). These observations suggest the involvement of
cAMP production in these immunoregulatory effects by histamine.
|
To prove that the effects of histamine are not limited to LPS stimulation,
PBMC were stimulated with SAC, in the presence or absence of increasing
concentrations of histamine. As shown in Figure 6![]() ,
A and B, histamine potently suppressed IL-12 (p70)
production in a concentration-dependent manner, while it potentiated
the production of IL-10. Similar to the whole blood assay, the
effects of histamine on SAC-induced IL-12 and IL-10 production were
blocked by the H2 receptor antagonist cimetidine, implicating once
again the role of H2 receptors.
,
A and B, histamine potently suppressed IL-12 (p70)
production in a concentration-dependent manner, while it potentiated
the production of IL-10. Similar to the whole blood assay, the
effects of histamine on SAC-induced IL-12 and IL-10 production were
blocked by the H2 receptor antagonist cimetidine, implicating once
again the role of H2 receptors.
|
As shown in Figure 7![]() ,
A and C, PBL, free of monocytes, produced negligible
levels of IL-12 (p40) and no detectable levels of IL-10 in response
to LPS. While PBMC produced significantly less IL-12 and IL-10 than
enriched monocytes, coculture of PBMC with monocytes resulted in
intermediate production of these two cytokines. These results
indicate that monocytes are the primary source of IL-12 and IL-10 in
the LPS-stimulated whole blood assay. This is in accordance with
previous results of others (20). To verify that
peripheral blood monocytes were directly affected by histamine,
enriched human monocytes were stimulated with LPS in the presence or
absence of increasing concentrations of histamine. As shown in Figure
7
,
A and C, PBL, free of monocytes, produced negligible
levels of IL-12 (p40) and no detectable levels of IL-10 in response
to LPS. While PBMC produced significantly less IL-12 and IL-10 than
enriched monocytes, coculture of PBMC with monocytes resulted in
intermediate production of these two cytokines. These results
indicate that monocytes are the primary source of IL-12 and IL-10 in
the LPS-stimulated whole blood assay. This is in accordance with
previous results of others (20). To verify that
peripheral blood monocytes were directly affected by histamine,
enriched human monocytes were stimulated with LPS in the presence or
absence of increasing concentrations of histamine. As shown in Figure
7![]() ,
B and D, histamine, as in the case of LPS-stimulated
whole blood or SAC-stimulated PBMC, substantially inhibited
LPS-induced IL-12 (p40) production in a concentration-dependent
manner, while it potentiated the production of IL-10 from human
monocytes. Similar to the findings with whole blood and PBMC, the
effect of histamine on LPS-induced IL-12 and IL-10 production from
monocytes was blocked by the H2 receptor antagonist cimetidine, thus
implicating the critical role of H2 receptors on monocytes as
mediating the effect of histamine in human peripheral whole blood or
PBMC. Since isolated, unprimed blood monocytes produce little IL-12
p70 when stimulated with LPS or SAC, the effect of histamine on p70
could not be reliably evaluated in these experiments.
,
B and D, histamine, as in the case of LPS-stimulated
whole blood or SAC-stimulated PBMC, substantially inhibited
LPS-induced IL-12 (p40) production in a concentration-dependent
manner, while it potentiated the production of IL-10 from human
monocytes. Similar to the findings with whole blood and PBMC, the
effect of histamine on LPS-induced IL-12 and IL-10 production from
monocytes was blocked by the H2 receptor antagonist cimetidine, thus
implicating the critical role of H2 receptors on monocytes as
mediating the effect of histamine in human peripheral whole blood or
PBMC. Since isolated, unprimed blood monocytes produce little IL-12
p70 when stimulated with LPS or SAC, the effect of histamine on p70
could not be reliably evaluated in these experiments.
|
| Discussion |
|---|
 .
Recent evidence, however, indicates that factors other than cytokines
have important regulatory influences on IL-12 and IL-10 production.
These factors include the inflammatory mediators PGE2 (20)
and nitric oxide (21), and the two major stress
hormones, glucocorticoids and catecholamines (22).
.
Recent evidence, however, indicates that factors other than cytokines
have important regulatory influences on IL-12 and IL-10 production.
These factors include the inflammatory mediators PGE2 (20)
and nitric oxide (21), and the two major stress
hormones, glucocorticoids and catecholamines (22).
Here we report that histamine mediates dose-dependent inhibition of human bioactive IL-12 and stimulation of IL-10 production in whole blood cultures stimulated with LPS. Cimetidine, an H2 receptor antagonist, blocked the effects of histamine on IL-12 and IL-10 production, while H1 and H3 receptor antagonists failed to alter the effects of histamine. These data suggest that histamine is modulating the production of these two cytokines via the H2 receptor. This conclusion is further substantiated by the finding that dimaprit, an H2 receptor agonist, mimicked dose-dependently the effect of histamine. In agreement with previous studies (20), we provide evidence that monocytes are the main IL-12- and IL-10-producing cells in LPS-stimulated human peripheral blood. Human monocytes and monocytic lines express H2 receptors, and stimulation of these receptors by histamine or dimaprit results in increased cAMP formation (23). In addition, we demonstrated that, in monocytes, the effects of histamine on IL-12 and IL-10 production were similar to those in whole blood and also were blocked by the H2 receptor antagonist cimetidine, while Ro 20–1724, a phosphodiesterase inhibitor, potentiated the effects of histamine on both IL-12 and IL-10 secretion. Thus, H2 receptors on monocytes appear to mediate the effects of histamine on IL-12 and IL-10 cytokine production in peripheral blood via an increase of intracellular cAMP. This is in accordance with recent data showing that increased cAMP induced by other mediators, such as PGE2 and catecholamines, have similar differential effects on IL-12 and IL-10 production (20, 22).
Monocytes/macrophages and other phagocytic cells are components of innate
or natural immune mechanisms, while lymphocytes are components of
acquired (adaptive) or specific immune mechanisms. Innate and specific
immunity, however, are not autonomous. Cells of the innate immune
system can determine which Ags the acquired immune system responds to
and the nature of that response (24). This can be
mediated through cytokines released by monocytes/macrophages that
play a critical role in influencing the Th1/Th2 pattern that in turn
dictates the type of immune response generated. Thus, IL-12 is a
central inducer of Th1 differentiation and serves as a bridge between
the innate and specific immunity (25), whereas
IL-10 antagonizes the activities of IL-12. The Th1/Th2 pattern is
often regarded as a balance between Th1/Th2 cell cytokine activities,
but our observations suggest that conditions related to increased
local or systemic levels of histamine may also contribute to Th1/Th2
balance. In particular, Th2-associated humoral immunity is
potentiated. Our data are consistent with previous studies showing
that histamine via H2 receptors inhibits TNF- production from monocytes and mast cells (26, 27),
while it potentiates IL-6 production from endothelial cells (28).
This view is further substantiated by a recent study demonstrating
that histamine, also via H2 receptors, inhibits IFN-
production by Th1-like cells but has no effect on IL-4 production
from Th2 clones (29). Moreover, histamine directly
enhances the production of human IgE from B cells (30),
while it inhibits T cell cytotoxicity. Thus, available data suggest
that histamine induces a Th2 shift at the level of monocytes, at the
level of Th cells, and directly on effector cells. Interestingly, in
the 1980s Rocklin and coworkers published several studies showing
that histamine induced suppressor-cell activity and suppressor factor
that were dependent of the presence of monocytes (31).
We can speculate that this factor might have been IL-10. In summary,
our data, considered in the context of these other studies, strongly
suggest that histamine, apart from exerting potent effector functions
in inflammation and allergy (mainly via H1 receptors), may have
important immunoregulatory functions via H2 receptors expressed on
immune cells.
production from monocytes and mast cells (26, 27),
while it potentiates IL-6 production from endothelial cells (28).
This view is further substantiated by a recent study demonstrating
that histamine, also via H2 receptors, inhibits IFN-
production by Th1-like cells but has no effect on IL-4 production
from Th2 clones (29). Moreover, histamine directly
enhances the production of human IgE from B cells (30),
while it inhibits T cell cytotoxicity. Thus, available data suggest
that histamine induces a Th2 shift at the level of monocytes, at the
level of Th cells, and directly on effector cells. Interestingly, in
the 1980s Rocklin and coworkers published several studies showing
that histamine induced suppressor-cell activity and suppressor factor
that were dependent of the presence of monocytes (31).
We can speculate that this factor might have been IL-10. In summary,
our data, considered in the context of these other studies, strongly
suggest that histamine, apart from exerting potent effector functions
in inflammation and allergy (mainly via H1 receptors), may have
important immunoregulatory functions via H2 receptors expressed on
immune cells.
The pattern of modulation exerted by histamine on IL-12 and IL-10 production might also be relevant to understanding host resistance to various infectious agents, where the selection of Th1- vs Th2- responses plays an important role. For example, recent evidence indicates that Th1 responses are important in the defense against infection with H. pylori (15). Now it is clear that this infection is the most common cause of chronic gastritis that in some cases progresses to peptic ulcer disease. An increased local concentration of histamine, induced by inflammatory or stress-related mediators, may play a pathogenic role in these conditions. It is tempting to speculate that a Th2 shift, induced by local excessive histamine production, may represent an additional pathogenic factor that participates in the development or extension of H. pylori infection. Our results also raise the question as to whether H2 receptor antagonists, widely used to prevent histamine-induced acid secretion by preventing the effect of histamine on cytokine-producing resident gastric monocytes/macrophages, may also favorably alter the host response to H. pylori infection and prevent progression into peptic ulcer disease, by restoring local Th1 responses.
It was recently reported that treatment with cimetidine correlated with increased survival in patients with gastric and colorectal cancer (17, 18). Histamine has been suggested to be a receptor-dependent "growth factor" in some colon, gastric, breast, and melanoma cell lines. Also, high concentrations of histamine have been measured within colorectal and breast cancer tissues, and large numbers of mast cells have been identified within tumor tissues (16). The above mentioned beneficial effect of cimetidine on survival from cancer remains poorly understood (18). Our results suggest that, by preventing the effect of histamine on IL-12 and IL-10 production, cimetidine may contribute to a restoration of Th1 responses and a better defense against these particular tumors.
Atopic reactions are characterized by dominant Th2 responses. Our results suggest that histamine might participate in a positive feedback loop, whereby allergen/Ag-IgE-induced release of histamine directly modulates IL-12 and IL-10 production that promotes and sustains a shift to IgE production.
In conclusion, we have demonstrated that histamine alters substantially the pattern of monocyte IL-12/IL-10 production and that this effect can be blocked by an H2 antagonist. Our results suggest that H2 receptor antagonists might be candidates for pharmacologic enhancement of Th1 functions in certain types of infections and tumors and for inhibiting a Th2 shift in certain allergic reactions.
| Acknowledgments |
|---|
| Footnotes |
|---|
2 Abbreviations used in this paper: Tc, T-cytotoxic; NOD,
nonobese diabetic; SAC, Staphylococcus aureus Cowan strain 1;
ND50, 50% neutralizing dose. ![]()
Received for publication November 3, 1997. Accepted for publication May 4, 1998.
| References |
|---|
release by mast cells through H2 and H3 receptors. Am.
J. Respir. Cell Mol. Biol. 14:620.
[Abstract]
)
release according to the phenotype of human Th0, Th1 and Th2 clones. Clin.
Exp. Immunol. 108:545.
[Medline]
Cimetidine enhances the hepatoprotective action of N-acetylcysteine in mice treated with toxic doses of paracetamol. Al-Mustafa ZH; Al-Ali AK; Qaw FS; Abdul-Cader Z Department of Pharmacology, College of Medicine and Medical Sciences, King Faisal University, Dammam, Saudi Arabia. Toxicology (IRELAND) Sep 5 1997, 121 (3) p223-8
Paracetamol, in toxic doses, is associated with extensive liver damage. This represents one of the common causes of morbidity and mortality in drug poisoning cases. This study was undertaken to investigate the possible potentiation of the hepatoprotective action of N-acetylcysteine (NAC) by cimetidine (CMD), an inhibitor of hepatic microsomal oxidative enzymes. The effects of NAC, cimetidine and the two in combination, administered 2 h post-paracetamol dose, on mortality, plasma glutamic oxaloacetic (GOT) and glutamic pyruvic (GPT) transaminase activities and hepatic reduced glutathione (GSH) levels were investigated in mice 24 h after treatment with a single oral dose of paracetamol (400 mg/kg). Both NAC and cimetidine caused a partial improvement of survival rate, plasma GOT and GPT activities. In addition, they prevented the depletion of hepatic GSH contents. However, concomitant administration of NAC and cimetidine produced a 100% survival rate and a marked reduction in plasma GOT and GPT activities to within the normal range, while significantly raising hepatic GSH concentrations to values close to those measured in saline-treated control animals. It is therefore concluded that cimetidine and N-acetylcysteine may have an additive hepatoprotective action in the treatment of paracetamol overdose.
Cimetidine protects against acetaminophen toxicity Jackson J.E. Sect. Clin. Pharmacol., Dept. Pharmacol., Univ. Arizona Health Sci. Cent., Tucson, AZ 85724 United States Life Sciences ( LIFE SCI. ) (United Kingdom) 1982, 31/1 (31-35)
Generally, acetaminophen (APAP) overdoses with elimination half-lives over 4 hr. sustain liver damage. In the following cases, cimetidine (C) seems to have protected against APAP toxicity. An 18 yr. old, 64 kg female smoker presented 6 hr. after taking 10 g APAP, 1200+ mg C, and small amounts of flurazepam and Sleepeze (methaprilene + scopolamine). Three plasma APAP levels (by HPLC) revealed an elimination half-life of 4.4 hr. C did not interfere with the APAP assay. Despite the long half-life in a patient with microsomal enzymes induced by smoking, no evidence of hepatotoxicity developed. A month later, the same patient overdosed with APAP alone. Three plasma levels revealed a 3.3 hr. half-life. Lack of toxicity in the presence of a long elimination half-life may indicate a protective action of C in APAP overdoses. To test the hypothesis that C protects against APAP heptatotoxicity, adult male Swiss mice were treated with 1200 mg/kg APAP by gavage. One hr. after APAP, the mice were treated with either 100 mg/kg C(n=19), 600 mg/kg N-acetylcysteine (standard therapy, n=17), or saline (control, n=17). Seven day survival was 24% among the controls, 68% with cimetidine treatment (v. control p <.05), and 100% with N-acetylcysteine (v. control P <.01; v. C, p>0.2). C inhibits the microsomal mono-oxygenase (cytochrome P-450) system which mediates acetaminophen hepatotoxicity. By preventing the formation of the toxic metabolites, C may prove useful in treating APAP overdosage. However, in our animal model it seems less effective than N-acetylcysteine.
END